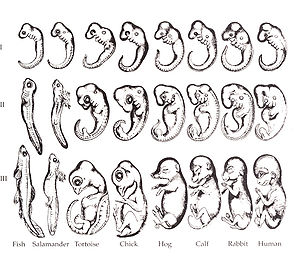
A copy of Haeckel's original drawings. Wikimedia
Note: This is the first of a series of articles (how long the 'series' will be depends on how long I can sustain it, of course) on Evo-Devo. I'm going to introduce Evo-Devo in this one, and then go over some interesting new research in the next few. I'm sorry if it seems like I'm only good for Evolution articles. I think it's true.
Here’s two technical words that if mentioned in conversation are likely to make your non-science friends glaze over: ontogeny and phylogeny. Neither of them is a hard concept, but perhaps because of the confusing repetition of the –geny’s, its hard to remember which is which. A similar quandary: even after three years of Biology courses, I sometimes find myself needing to Google which is which from paralogs and orthologs.
Ontogeny essentially refers to the mode of development of an organism, all the way from the first cell (the zygote) which resulted due to fusion of the maternal and paternal gametes, to the mature adult form. That’s simple enough. Phylogeny is the trickier one. Much like a color spectrum, phylogeny grades species based on how evolutionarily ‘similar’ they are to the species closest to them, and also based on the time point in life history at which they first appeared. Unlike a color spectrum, phylogeny is expressed in the form of trees, which of course can express the idea that species can sometimes be ‘side-branches’ and not, strictly speaking, a point on the continuum.
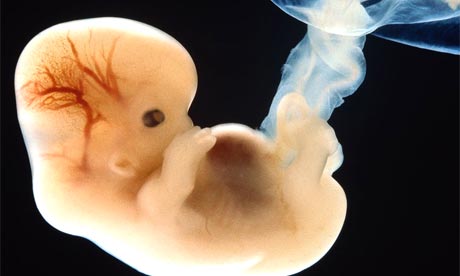
Yes, embryos are generally rather strange-looking.
Now that we know about ontogeny and phylogeny, we can consider the principal connection between them. This is the idea that ‘ontogeny recapitulates phylogeny’. Recapitulation theory, first put forth (most controversially) by Ernst Haeckel, states that the embryo form of an organism reveals a lot of information about the organism’s evolutionary history. Thus, if we look at the forms that the embryo takes at many points in its development, we’ll see some structures which develop all the way to the adult and some which don’t. And all of these structures will be indicative of structures that are either present in the organism or in the organism’s ancestors that have long since disappeared. This connection between ontogeny and phylogeny brings together two disparate disciplines: Evolutionary Biology and Developmental Biology (colloquially and often academically called Evo-Devo).
Strictly speaking, recapitulation theory is a discredited one because embryonic structures of more recently evolved organisms do not necessarily reflect structures present in its remote ancestors. The reason this is all still relevant, however, is that ontogeny and phylogeny are still very much related, as Stephen Jay Gould theorized in his opus Ontogeny and Phylogeny. The central focus today is that it’s not just structures in the embryo that reveal a great deal about phylogenetic history. It’s also the molecules involved in making things happen for the embryo, and the genes which are involved in making those molecules. And when this information is compared across many species, a startling truth emerges: almost across the board, a surprisingly small set of genes is utilized again and again in species to do similar things. For instance, the gene Pax6 has been found to govern eye development without exception across the staggering array of vertebrates and arthropods. A lack of Pax6 had previously been known to be associated with aniridia, a form of blindness where the retina of the eye develops imperfectly. And here’s the kicker: if a blind mouse embryo lacking Pax6 is transfected with the Pax6 gene from the fruit fly Drosophila, eye development is ‘rescued’. The blind mouse miraculously developed healthy mouse eyes! Pax 6, as a result, is one of a few genes that is so important as to be conserved over vast evolutionary distances and still be utilized for the same function. This limited repertoire of genes, unsurprisingly, foreshadows many perennial research foci in Evo-Devo.
I would really like to go into the specifics of a few open questions in the field, particularly concerning how processes like axis patterning and germ cell development are highly conserved across many species. This will require a whole other blog post, however. Till then, you may as well amuse yourself with the embryo image above.
For an interview of one of the leading Evo-Devo researchers, Dr. Cliff Tabin (who happens to be working on Darwin's finches) click here.
Kamil
Strictly speaking, recapitulation theory is a discredited one because embryonic structures of more recently evolved organisms do not necessarily reflect structures present in its remote ancestors. The reason this is all still relevant, however, is that ontogeny and phylogeny are still very much related, as Stephen Jay Gould theorized in his opus Ontogeny and Phylogeny. The central focus today is that it’s not just structures in the embryo that reveal a great deal about phylogenetic history. It’s also the molecules involved in making things happen for the embryo, and the genes which are involved in making those molecules. And when this information is compared across many species, a startling truth emerges: almost across the board, a surprisingly small set of genes is utilized again and again in species to do similar things. For instance, the gene Pax6 has been found to govern eye development without exception across the staggering array of vertebrates and arthropods. A lack of Pax6 had previously been known to be associated with aniridia, a form of blindness where the retina of the eye develops imperfectly. And here’s the kicker: if a blind mouse embryo lacking Pax6 is transfected with the Pax6 gene from the fruit fly Drosophila, eye development is ‘rescued’. The blind mouse miraculously developed healthy mouse eyes! Pax 6, as a result, is one of a few genes that is so important as to be conserved over vast evolutionary distances and still be utilized for the same function. This limited repertoire of genes, unsurprisingly, foreshadows many perennial research foci in Evo-Devo.
I would really like to go into the specifics of a few open questions in the field, particularly concerning how processes like axis patterning and germ cell development are highly conserved across many species. This will require a whole other blog post, however. Till then, you may as well amuse yourself with the embryo image above.
For an interview of one of the leading Evo-Devo researchers, Dr. Cliff Tabin (who happens to be working on Darwin's finches) click here.
Kamil
 RSS Feed
RSS Feed